ojo
El ojo comenzó como un punto en la epidermis capaz de la detección de luz.
Evolución de los ojos y fotorreceptores.
Estoy escribiendo una breve semblanza del ojo humano y la luz; concentrándome en el ojo como receptor de esta. La finalidad es allanar el camino para la creación de una pieza centrada en la luz y el ojo. En adelante hablaré del ojo como un correlativo de la luz.
Luz y ojos para ver
Según Dan-E Nilsson en su artículo La evolución del ojo y su base funcional, la evolución del ojo pasó por cuatro momentos: Fotorrecepción no direccional; fotorrecepción direccional; visión de baja resolución; y visión de alta resolución. Sigue diciendo Nilsson: El proceso completo, desde la primera opsina hasta la visión de alta resolución, tardó unos 170 millones de años y se completó en gran medida para el inicio del Cámbrico, hace unos 530 millones de años.
Del mismo modo, dentro de esta concepción evolutiva, que considera que el ojo evolucionó porque es un órgano que asegura la supervivencia y reproducción de una especie, se encuentran Mario Eduardo Guido y Pedro Panzetta, quienes afirman: No sólo los ojos complejos son útiles; de hecho muchos animales poseen ojos menos complejos o estructuras simples que detectan luz; estos 'ojos simples' son óptimos para asegurar la supervivencia y reproducción de los organismos.
Un punto de vista similar se encuentra en El origen del Ojo: Cómo la Biología Evolutiva desafía al Diseño Inteligente, en donde se indica lo siguiente:
- La aparición del órgano visual primitivo representó una tremenda ventaja evolutiva, permitiendo a los animales convertirse en cazadores activos.
- El ojo ha evolucionado a partir de estructuras simples con funciones limitadas hasta convertirse en órganos más complejos capaces de percibir la visión tridimensional y en color.
- El ojo ha evolucionado desde rudimentarias zonas fotosensibles incapaces de formar imágenes hasta los sofisticados órganos que conocemos hoy.
- El ojo inicialmente evolucionó en un entorno acuático y nunca se adaptó para la vida en ambientes aéreo.
Clasificación del comportamiento controlado por fotorreceptores, de Nilsson
- Clase I: Fotorrecepción no direccional: Con solo monitorear la intensidad de la luz ambiental, es posible obtener información para una serie de comportamientos importantes, como activar o desactivar comportamientos directamente en relación con la hora del día o indirectamente al proporcionar información a un reloj circadiano.
- Clase II: Fotorrecepción direccional: Si el fotorreceptor está protegido de tal manera que detecta la luz en algunas direcciones, pero no en otras, es muy adecuado para determinar la dirección hacia o lejos de la luz en un animal en movimiento. Un comportamiento obvio guiado por la fotorrecepción direccional es la fototaxis.
- Clase III: Visión de baja resolución: Gracias a una matriz de fotorreceptores direccionales, se pueden realizar lecturas simultáneas de la luminancia en diferentes direcciones. Se utilizan para controlar la velocidad y la dirección de la locomoción, así como la trayectoria en relación con las estructuras circundantes; así como en las respuestas de evitación de objetos (anticolisión). Esta clase introduce la imagen y la resolución espacial, por tanto, representa los primeros ojos verdaderos y el origen de la visión, más allá de la simple fotorrecepción.
- Clase IV: Visión de alta resolución: Con solo aumentar la resolución espacial, es posible participar en la detección, persecución y comunicación con otros animales.
Requisitos fundamentales de rendimiento para las cuatro clases, de Nilsson
Se trata de las exigencias de tiempo de integración (velocidad de respuesta), ángulo de detección y precisión de detección.
- Tiempo de integración: Claramente, el tiempo de integración necesario para evitar el desenfoque de movimiento en la visión de alta resolución es mucho más corto que el necesario para monitorear el ciclo de luz diario. La sensibilidad a la luz no direccional necesaria para las tareas de clase I se basa en la variación temporal en la radiancia de la fuente de luz principal, como el sol o la luna, y esta variación es típicamente lenta. Las mediciones experimentales del tiempo de integración en fotorreceptores no direccionales están en el rango de 10 a 30 minutos; mientras que para la fotorrecepción direccional y la visión verdadera, el movimiento propio de los animales también provocará cambios de señal mucho más rápidos, lo que requiere tiempos de integración de 1 segundo o menos.
- Ángulo de detección: El ángulo de detección requerido se estrecha para cada clase superior. Por definición, la fotorrecepción no direccional (clase I) no debe estar restringida en ángulo. Las tareas de clase II requieren direccionalidad, como la fototaxis. Los fotorreceptores para la fototaxis suelen captar luz en un rango de 100° a 180°. Las clases III y IV requieren ángulos de detección mucho más pequeños porque estos determinarán directamente la agudeza visual, que varía desde 30° hasta fracciones de grado.
- Precisión de detección: Los cambios de radiancia entre un día soleado y una noche estrellada abarcan aproximadamente 8 unidades logarítmicas. Incluso con una precisión de tan solo el 30%, es posible detectar con fiabilidad el inicio del anochecer y el amanecer.
Las tareas de clase I pueden realizarse a intensidades inferiores a la luz de las estrellas con células sin ayuda y morfológicamente no especializadas. Las tareas de clase II también pueden ser realizadas por dichas células receptoras simples, pero solo hasta intensidades de medio crepúsculo. Para la operación a intensidades más bajas, es necesario introducir apilamiento de membranas, que es un prerrequisito para que las tareas de clase III funcionen incluso a intensidades de luz diurna. Para extender las tareas de clase III por debajo de las intensidades de medio crepúsculo, es necesario agregar óptica de enfoque. La visión de alta resolución requiere tanto apilamiento de membranas como óptica de enfoque para todas las intensidades, y las aperturas suficientemente grandes permiten la operación hasta intensidades nocturnas.
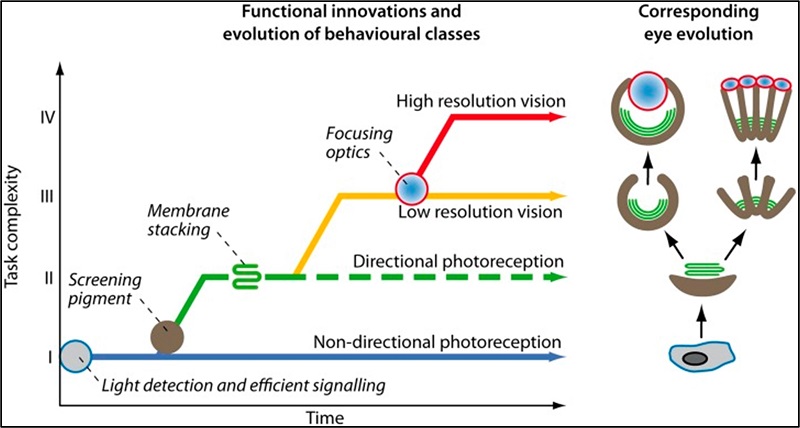
Ilustración esquemática de la evolución de los comportamientos fotorreceptivos en una escala vertical de complejidad de la tarea, indicándose la posición de las principales innovaciones funcionales.
Lo que sí es evidente es que primero fue la luz y luego el ojo, como afirma Nilsson: Uno de los primeros pasos en la evolución del ojo debe haber sido la aparición de una reacción química dependiente de la luz acoplada a un sistema de señalización.
Máquina visual
Ahora bien, los humanos tenemos visión en alta resolución, pero no vemos en la oscuridad total; sin embargo, en ciertas condiciones nocturnas podemos ver gracias a la apertura de la pupila. Actualmente, esta condición está superada gracias a la luz generada con energía eléctrica.
En general se considera al último cuarto del siglo XIX como el momento en el que la energía eléctrica comenzó a producir luz. En México, la Comisión Federal de Electricidad dice que la primera planta generadora que se instaló en el país (1879) estuvo en León, Guanajuato, y era utilizada por la fábrica textil ‘La Americana’. Y que para el final del gobierno de Porfirio Díaz ya se contaba con alumbrado público en la Plaza de la Constitución, la Alameda y la calle Reforma.
Sin profundizar en esta historia, puedo decir que apenas han pasado ciento cincuenta años en los que los humanos hemos mejorado nuestra visión nocturna de manera artificial, gracias a la luz eléctrica. Este dato no es menor, ya que como lo señalan las referencias, la evolución del ojo tiene al menos 700 millones de años y sucedió de manera natural, es decir, sin intervención de tecnologías.
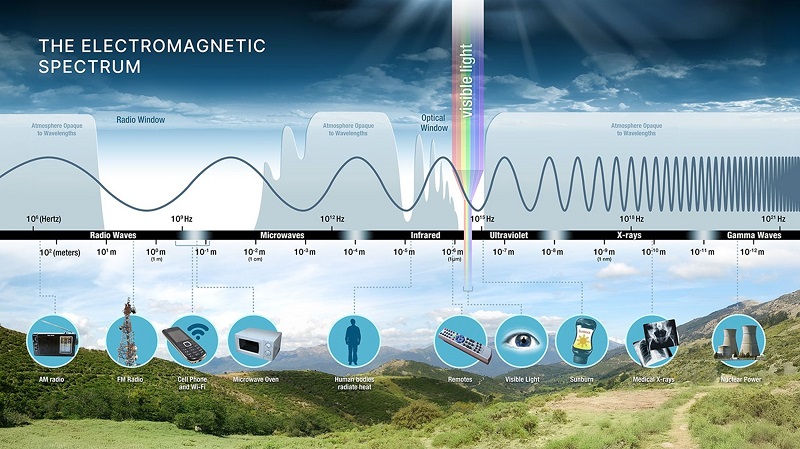
Aunado a lo anterior, aunque tenemos luz artificial nocturna, aún no captamos buena parte del espectro electromagnético. El ojo humano solo puede detectar una pequeña parte del espectro, es lo que llamamos ‘luz visible’. Las radios captan otra porción del espectro y lo mismo sucede con las máquinas de rayos X.